Article Text

Abstract
Background—Laminin receptors are involved in cell-extracellular matrix interactions in malignant cells that show invasion and metastasis. Hepatocellular carcinoma frequently shows early invasion into blood vessels, and intrahepatic and extrahepatic metastases. However, the role of laminin receptors in hepatocellular carcinoma is unknown.
Aims—To examine the expression of mRNA for laminin receptors and their isoforms in hepatocellular carcinoma.
Methods—The expression of several laminin receptors, including α1 integrin, α6 integrin and its isoforms α6A and α6B, β1 integrin and its isoforms β1A and β1B, and 32kD/67kDa laminin binding protein was examined in human hepatocellular carcinomas and non-cancerous liver tissues using the reverse transcription polymerase chain reaction.
Results—α6 Integrin, β1 integrin, and laminin binding protein showed notably increased expression in hepatocellular carcinoma, compared with non-cancerous liver tissue, although the α1 integrin did not show a significant change. Furthermore, β1B integrin, a splicing variant of β1 integrin, was overexpressed in hepatocellular carcinoma while the β1A integrin isoform did not show significant changes between hepatocellular carcinoma and surrounding non-cancerous liver tissue.
Conclusions—The differential upregulation of laminin receptors and their splicing isoforms was shown in hepatocellular carcinoma, suggesting that certain laminin receptors and their isoforms may be involved in the development and progression of hepatocellular carcinoma.
- laminin receptor
- integrin α6β1
- hepatocellular carcinoma
Statistics from Altmetric.com
The process of tumour development and metastasis appears to involve alterations in cell-cell and/or cell-extracellular matrix (ECM) interactions mediated by specific receptors. The interaction between transformed cells and the basement membrane is an important step in the development of invasion and metastasis.1 Laminin, one of the major components of the basement membrane, was first purified from tumour cells as a heterotrimeric protein consisting of A, B1, and B2 subunits,2 later renamed α1, β1, and γ1.3 Recently homologues of laminin subunits have been cloned and revealed a variety of heterogeneity of laminins. To date, at least 11 distinct laminin isoforms have been identified.3 ,4 It has been reported that certain fragments of laminin inhibit the metastatic activity of some malignant cells,5 ,6 and the other fragments enhance the metastatic activity of tumour cells.7 These effects are thought to be mediated by specific laminin receptor(s). Therefore it may be useful to examine the expression of laminin receptors on malignant cells, to understand better their role in carcinogenesis, invasion, and metastasis.
Hepatocellular carcinoma (HCC) frequently shows invasion into blood vessels at an early stage as well as intrahepatic and extrahepatic metastases.8 The interaction between liver cancer cells and laminin through specific laminin receptors may be involved in the process of hepatocarcinogenesis. A variety of laminin receptors have been identified, including integrin which is a heterodimer composed of one α and one β subunit, and non-integrin families (reviewed in Mecham9). However, little data is available on the alterations of the ECM receptors in HCC, and their role in vascular invasion and metastasis. We examined the expression of several laminin receptor mRNAs, including α1 integrin, α6 integrin and its isoforms α6A and α6B, β1 integrin and its isoforms β1A and β1B, and 32/67 kDa laminin binding protein (LBP), in human HCC tissue using a semiquantitative reverse transcription polymerase chain reaction (RT-PCR) method. Here we show the usefulness of semiquantitative RT-PCR, and document the differential upregulation of laminin receptors and their splicing variants in human HCC.
Materials and methods
HUMAN LIVER SAMPLES
We examined tumour and corresponding non-cancerous liver tissue in 16 patients with HCC. There were two women and 14 men (mean (SD) age 60.7 (5.0) years, range 55–67). All 16 patients were positive for antihepatitis C virus antibody; four had chronic hepatitis and 12 liver cirrhosis diagnosed by histopathological examination under a light microscope. We also examined liver tissue from three subjects with normal liver histology.
The liver specimens were obtained by surgical resection performed for treatment of the hepatic neoplasms (eight patients), or by autopsy performed within three hours of death (eight patients) at the hospital of the Saga Medical School. A portion of each human liver specimen was used for histological diagnosis by light microscopy. The remaining liver tissue was immediately frozen in liquid nitrogen and stored at −80°C for RNA extraction. The histological grading of HCC was based on Edmondson’s grading system,10 and comprised two grade I, 11 grade II, and three grade III disease. The clinical stage was based on the TNM classification system of the UICC,11 and comprised two stage I, three stage II, one stage III, and nine stage IV disease.
RNA EXTRACTION AND CONFIRMATION OF INTEGRITY
Total RNA was extracted from liver tissue by using the guanidinium method as previously described.12 After RNA extraction, to confirm the integrity of total RNA extracted, especially from postmortem liver samples, 1 μg of each total RNA sample was electrophoresed in an agarose gel, stained with ethidium bromide, visualised under ultraviolet (UV) light, and compared with the total RNA extracted from surgical liver samples. Postmortem livers of 23 patients were subjected to total RNA extraction; only the RNA of eight patients, which did not show significant degradation of 28S and 18S ribosomal RNA, was used for the experiment. mRNA expression of several genes was compared between surgical liver samples and postmortem liver samples. Semiquantitative RT-PCR of oncogene Ets-1, one of the extracellular matrix components of the laminin α2 chain, and housekeeping gene GAPDH were performed, and results showed no significant differences in the expression of these genes between surgical samples and postmortem samples (data not shown).
OLIGONUCLEOTIDES
Table 1 lists the sequences of the oligonucleotide primers used for the RT-PCR reactions.
Oligonucleotide primers used for RT-PCR analysis
RT-PCR AND CONFIRMATION OF RT-PCR PRODUCTS
cDNA (complementary DNA) was synthesised from 1 μg of total RNA with or without reverse transcriptase with random primers (Takara, Kyoto). The reaction mixture was then subjected to PCR amplification with specific forward and reverse oligonucleotide primers for 40 cycles consisting of heat denaturation (94°C for 1.5 minutes), annealing (50°C for 1.5 minutes), and extension (72°C for two minutes). PCR products were size fractionated on 2% agarose gels, and visualised under UV light. Aliquots of each RT-PCR product were used for digestion with specific restriction enzymes to confirm the sequences of the products: α1 with HinfI, α6 with XhoI, β1 with BglII, LBP with PstI, and β-actin with TaqI. All of the restriction enzymes used were purchased from Takara.
SEMIQUANTITATIVE RT-PCR
cDNA was synthesised from 1 × 10−5 to 1 μg of total RNA with reverse transcriptase (BRL, Gaithersburg, Maryland, USA) using random primers (Takara). Subsequently, the reverse transcriptase reaction mixture was used for PCR amplification with forward and reverse oligonucleotide primers, using a PCR kit with Taq DNA polymerase (Perkin Elmer/Cetus, Norwalk, Connecticut, USA). Each PCR reaction mixture was subjected to 15–40 cycles consisting of heat denaturation (94°C for 1.5 minutes), annealing (50°C for 1.5 minutes), and extension (72°C for two minutes). The PCR products were size fractionated on 2% agarose gels, and visualised under UV light. The PCR products were quantified using a negative film of PCR products with densitometric scanning. Isoform specific primers reported previously were used for the detection of the α6 and β1 isoforms (table 1). Reverse transcription was performed from 1 μg of total RNA, and 30 cycles of PCR were performed.
STATISTICAL ANALYSIS
The Dunnet test was used for non-parametric multiple comparisons of RT-PCR products among the normal controls, chronically diseased livers, and hepatocellular carcinomas.
Results
DETECTION AND CONFIRMATION OF THE RT-PCR PRODUCTS
To confirm that the RT-PCR products were RNA dependent, PCR was performed with or without reverse transcription (fig 1, lanes 1 and 2 of each RT-PCR). All the samples without reverse transcription showed no PCR products, indicating that these products were RNA dependent. To identify the sequences of the RT-PCR products, these products were digested with specific restriction enzymes, based on their sequences (table 1). Each reaction with specific restriction enzymes showed fragments of the predicted size (fig 1, lane 3 of each RT-PCR).
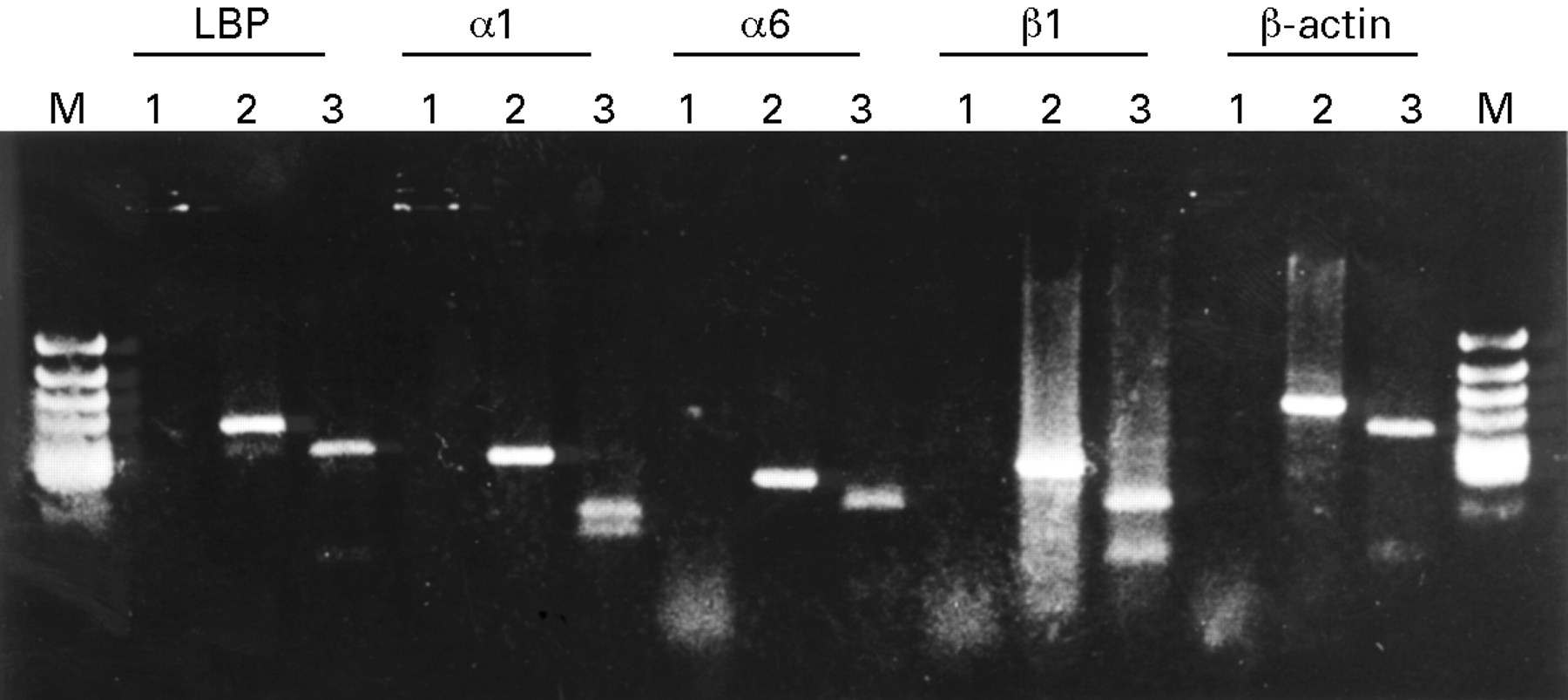
RT-PCR products of laminin binding protein (LBP), α1 integrin, α6 integrin, β1 integrin, and β-actin separated by 2% agarose gel electrophoresis. Lane 1: RT-PCR without reverse transcriptase. Lane 2: RT-PCR with reverse transcriptase. Lane 3: the products of lane 2 digested with a specific restriction enzyme based on the sequence data.
SEMIQUANTIFICATION OF RT-PCR
We first determined that the intensity of the signals obtained by this RT-PCR method was dependent on both the amount of RNA present, and on the number of cycles of PCR.20 We performed reverse transcription from serially diluted RNA samples, 1 to 1 × 10−5 μg, and subsequently subjected these samples to PCR for 30 cycles. Following agarose gel electrophoresis and staining with ethidium bromide, the intensities of the visualised PCR products were evaluated by densitometric scanning (fig 2A). Results showed that the signal increased exponentially according to the amount of total RNA, except for β-actin, which reached a plateau at 1 × 10−1 μg of total RNA. Next we performed RT-PCR from 1 μg of total RNA while changing the number of PCR cycles from 15 to 40, to see changes in the signals according to the cycle number. The PCR products were visualised by ethidium bromide staining and UV light, and evaluated by densitometry (fig 2B). The signals of the RT-PCR products increased exponentially as the number of PCR cycles increased, although the signals showed a tendency to reach a plateau at more than 35 cycles. After working out these conditions, we examined the mRNA levels of the laminin receptors in human liver tissue using 1 μg of total RNA and 30 cycles of PCR. The expression of β-actin mRNA was used as a loading control, using 1 × 10−1 μg of total RNA and 30 cycles of PCR.

Densitometric semiquantification of the RT-PCR products showed association with the amount of RNA applied (A), and the number of PCR cycles (B).
EXPRESSION OF LAMININ RECEPTORS IN HUMAN HEPATOCELLULAR CARCINOMA
In normal liver tissue, α1 integrin and β1 integrin mRNA were expressed highly, and α6 integrin and laminin binding protein (LBP) mRNA were also expressed, but weakly (figs 3 and 4). In the non-cancerous liver tissue with chronic liver disease, β1 integrin and LBP showed increased expression compared with normal liver tissue, although α1 integrin and α6 integrin did not show a significant increase (figs 3 and 4). Furthermore, α6 integrin, β1 integrin, and LBP mRNAs are upregulated in HCC tissue compared with non-cancerous, chronically diseased liver tissue, although α1 integrin mRNA in HCC did not show a significant increase compared with non-cancerous liver tissue (figs 3 and 4). There are some extra bands larger than expected in the β1 integrin reaction in HCC samples in fig 3. Some were not reproducible, and some were sequenced directly, but no homology with the β1 integrin sequence was found (data not shown). Therefore, these products were considered to be non-specific mispriming products or as yet unknown gene products. When we compared the expression of two laminin receptors, α6 integrin and LBP, both of which showed increased expression in HCC tissue, α6 integrin showed a more prominent increase than did LBP. These data showed increased expression of some integrins and LBP in chronic viral liver disease, and showed further differential upregulation of α6 integrin, β1 integrin, and LBP in human HCC.
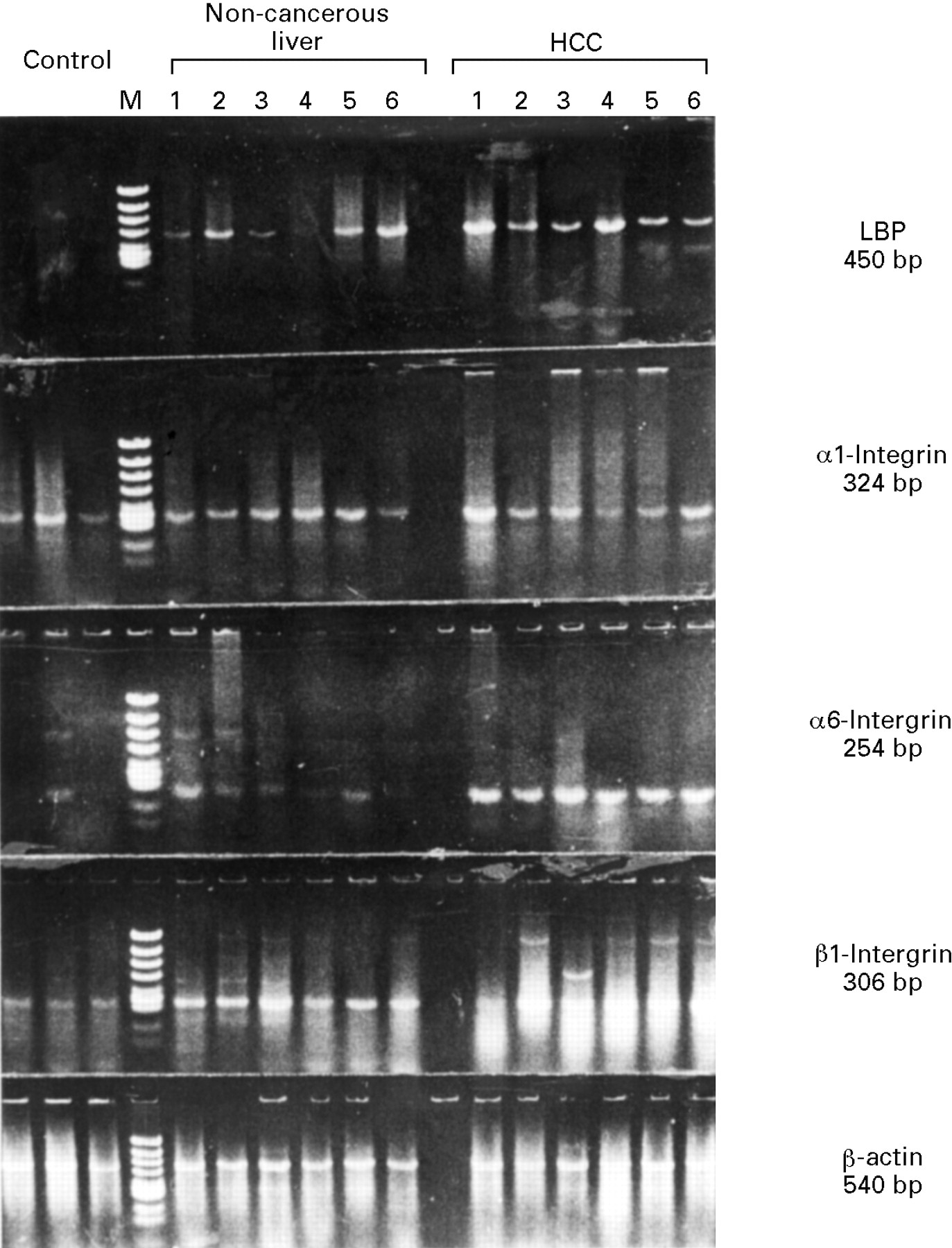
Gene expression of LBP, α1 integrin, α6 integrin, β1 integrin, and β-actin in six representative samples of human liver tissue detected by RT-PCR. Identical lane numbers indicate that the tissue was obtained from the same patient with chronic liver disease and HCC.

Semiquantitative analysis of LBP, α1 integrin, α6 integrin, β1 integrin, and β-actin gene expression in normal liver (N, n=3), chronically diseased, non-cancerous liver (L, n=16), and HCC tissue (n=16) detected by RT-PCR. Data are presented as mean (SD). *p<0.05; **p<0.01; *** p<0.001.
EXPRESSION OF ISOFORMS OF α6 AND β1 INTEGRIN
The alternative splicing derived isoforms of α6 integrin, α6A and α6B, and of β1 integrin, β1A and β1B, were amplified by RT-PCR with isoform specific primers.18 ,19 The same RT-PCR conditions used for the other integrins were used for the semiquantitative analysis of these isoforms. Although α6A and α6B mRNA were amplified by similar amounts in normal liver, much more α6B mRNA than α6A was amplified in chronically diseased, non-cancerous liver tissue and HCC tissue (fig 5A). The intensity of total α6 integrin expression was increased in HCC compared with normal liver or non-cancerous, chronically diseased liver (fig 3). When the ratio of α6B:α6A was compared among these groups, the amount of α6B mRNA expression in non-cancerous liver and HCC was significantly increased compared with normal liver (fig 5B). There was no difference in the α6B:α6A ratio between chronically diseased, non-cancerous liver and HCC.
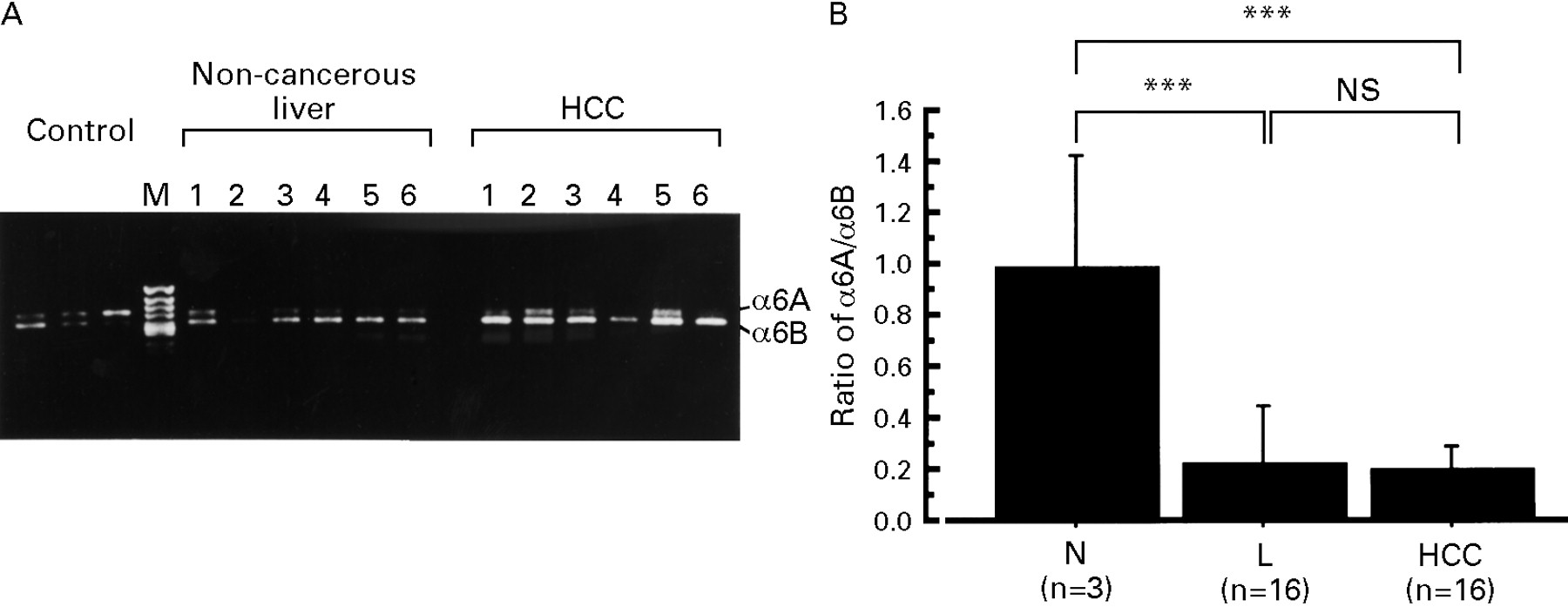
(A) Representative expression of the α6 integrin isoforms, α6A and α6B, by RT-PCR from six patients. (B) The α6B:α6A ratio detected by densitometry in normal liver (N, n=3), chronically diseased, non-cancerous liver (L, n=16), and HCC (n=16). Values are presented as mean (SD). ***p<0.001.
In the case of the β1 integrin isoforms, β1A and β1B, there was a significant difference in the expression pattern of these two isoforms (fig 6). The expression of β1B was significantly increased in HCC compared with non-cancerous liver tissue. Although the expression of β1A integrin in chronic liver disease and HCC was increased compared with normal liver, there was no significant difference between HCC and non-cancerous liver tissue.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Representative expression of the β1 integrin isoforms, β1A and β1B, by RT-PCR from six patients. (B) Semiquantification of β1A and β1B integrin in normal liver (N, n=3), chronically diseased non-cancerous liver (L, n=16), and HCC (n=16). Data are presented as mean (SD). *p<0.05; **p<0.01.
Discussion
Several laminin receptors including integrin have been reported to be present in normal and fibrotic liver.21 ,22 In normal liver, integrin α1 and β1 are expressed in hepatocyte and sinusoidal endothelial cells, although integrin α6 is only expressed in biliary cells and vascular endothelial cells. In inflammatory and fibrotic liver disease, α6 integrin showed increased expression in hepatocytes and sinusoidal endothelial cells. However, little is known about the expression of laminin receptors in liver tumours. Laminin receptor(s) are thought to be involved in a wide variety of biological events including development of tumour invasion and metastasis through cell-cell and/or cell-ECM interactions.1 ,5-7 ,9 Usually, the vascular structure in hepatocellular carcinoma contains basement membrane components, such as type IV collagen and laminin.23 ,24 Therefore, hepatocellular carcinoma cells may have some interaction with laminin, the major component of the basement membrane, through their specific laminin receptor(s). It has been reported that one of the laminin receptors, LBP, shows increased expression in HCC, and that this correlates with the histological grade.24 Recently, an immunohistochemical study showed de novo expression of several integrin molecules including α2, α3, and α6 integrin in HCC.25 More recently, overexpression of α6 integrin in human HCC has been reported.26 In the present study we investigated the mRNA levels of several laminin receptors in human HCC: LBP and two integrins, α1 and α6, both of which make heterodimers with the β1 integrin subunit. Furthermore, we also investigated the expression of two splicing variants of the cytoplasmic domain of α6 integrin, α6A and α6B, and of the cytoplasmic domain of β1 integrin, β1A and β1B. These variants are expressed in a tissue specific and cell type specific manner,18 ,19 ,27 ,28 accompany the changes in amino acid sequences of their cytoplasmic domains, and mediate different signals.29-31
We used RT-PCR to investigate the gene expression of these adhesion molecules because this method made it possible to detect the small amounts of mRNA obtained from the tiny tissue samples. For the semiquantitative assay of these mRNAs, it is necessary to show that the amount of RT-PCR product is dependent on both the amount of RNA and the number of PCR cycles.20 We showed that the amount of RT-PCR products detected by densitometry correlated with the amount of RNA present and with the number of PCR cycles.
Our data showed that the expression of LBP and β1 integrin was increased in fibrotic liver disease compared with normal liver tissue, and that the expression of LBP, α6 integrin, and β1 integrin was further increased in HCC compared with corresponding non-cancerous liver tissue. When we compared LBP and α6 integrin in HCC, the increase in α6 integrin expression was more prominent than that of LBP. These data suggest that the expression of laminin receptors is closely associated with the development and progression of HCC, and that they are differentially upregulated. It has been reported that in several malignant tumours including HCC, the expression of laminin receptors, including LBP and α6 integrin, correlates with invasiveness and metastatic activity,24 ,25 ,32-34 as well as with malignant transformation.35 Furthermore, our results on the expression of the isoforms of α6 integrin and β1 integrin show that the alternative splicing of these two integrins is also differentially regulated in non-cancerous liver tissue and HCC. Therefore, the altered splicing pattern of these integrins may contribute to the altered expression pattern of α6β1 integrin, and to the altered response of HCC cells to laminin because of the changes in the amino acid sequences of the cytoplasmic domain which mediates cellular-extracellular signalling.
In conclusion, this differential expression among laminin receptors and their isoforms in HCC may correspond to their different roles in cell-ECM interactions including the processes of malignant transformation, invasion, and metastasis. As most of our cases of HCC were advanced stage and had a similar histological grade, we could not show a relation between the expression of laminin receptors and the grade of the malignancy or disease progression, although we were able to show a close relation between the differential upregulation of laminin receptors and the development of HCC. Another point to be studied is which types of cells in HCC are responsible for the differential expression of these integrin molecules. To investigate the localisation and type of cells producing these integrin isoforms, immunohistochemical staining with specific antibodies or in situ hybridisation with specific probes, which can be analysed quantitatively, should be useful. Therefore, the expression and function of laminin receptors and their isoforms during hepatocarcinogenesis and tumour progression requires further study.